
Conduction of an impulse in human nerves relies upon the electrochemical activity of the individual neuron fibers inside the nerve. Each fiber (axon) inside the nerve is capable of propagating an action potential if the stimulus is strong enough to bring the membrane potential of the neuron to the threshold value. Depending on the strength of the stimulus, not all fibers in a nerve may fire. Despite this, the action potential is an all-or-none phenomenon, where it will always have the same amplitude and time course for a single neuron. When action potentials are recorded via recording electrodes on the skin, the result is a “compound action potential,” which is an algebraic sum of action potentials from many cells. We have already observed compound muscle action potentials (CMAP) in the EMGs of previous labs.
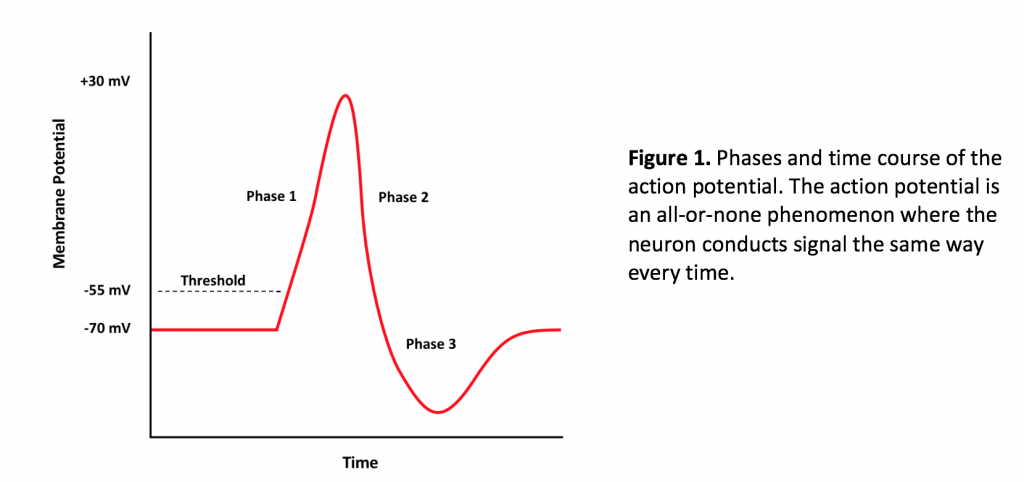
The action potential is the converse of the cell membrane resting potential, which, for neurons registers -70 mV on the voltmeter. The action potential involves several steps, starting with the onset of a stimulus. Most of the time, stimuli generating action potentials are chemical, where a ligand, a neurotransmitter, binds to a ligand-gated ion channel in the cell membrane of the dendrite or cell body of the neuron. If the graded potential is excitatory and sufficiently strong, the dissipation of charge across the cell body membrane reaches the axon hillock and depolarizes the cell membrane to -55 mV. The axon hillock is a region of high voltage-gated ion channel density. Voltage gated channels open in response to threshold depolarization of the membrane, leading to an influx of sodium (via the voltage gated sodium channel), bringing about the onset of Phase 1 of the action potential (Figure 1). The influx of sodium drives the membrane potential to +30 mV, which is the peak of the action potential on a graph. From here, voltage gated potassium channels, which slowly open near the end of Phase 1 of the action potential, allow efflux of potassium from the cell. The membrane repolarizes in response to the efflux of potassium, and the membrane potential reaches resting potential at the end of Phase 2. However, because the potassium channels close very slowly compared to sodium channels, the continued leakage of potassium from the cell results in membrane hyperpolarization, during which the membrane potential reaches -90 mV. At the end of phase 3, the activity of the sodium/potassium ATPase active transporter, which exports three sodium ions and imports two potassium ions helps to bring the membrane potential back to -70 mV (along with passive sodium and potassium leak channels). In order to communicate the signal of the action potential to another cell, synaptic transmission must occur. Synaptic transmission is the rate-limiting step of neural transmission, meaning that it is the slowest part of signal transmission. During synaptic transmission, the action potential reaches the axon termini of the pre-synaptic cell. The influx of sodium depolarizes the membrane of the termini and opens voltage gated calcium channels. Calcium enters the cell, resulting in the fusion of neurotransmitter-filled vesicles with the pre-synaptic membrane. Neurotransmitter diffuses across the synaptic cleft and binds to a receptor on the post-synaptic cell. The binding of neurotransmitter to a receptor will cause a post-synaptic potential (PSP) (a graded potential). Post-synaptic potentials may be excitatory or inhibitory, depending on the interaction of the neurotransmitter with the receptor.
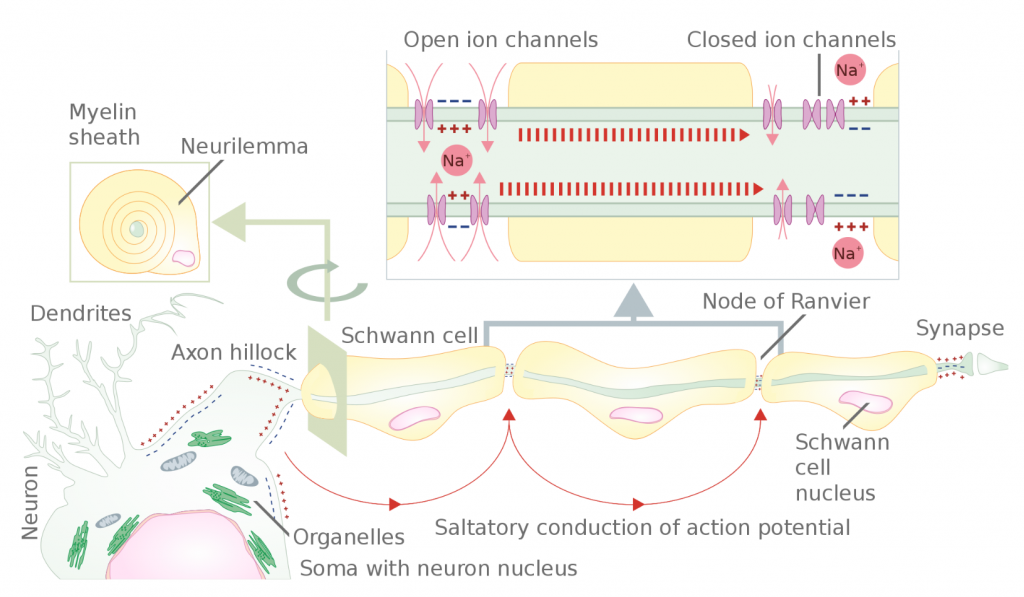
The speed of neuron transmission generally depends upon two factors: axon size and degree of myelination. Larger diameter axons transmit signal faster due to lower resistance to the flow of charge inside the axon. Myelinated axons (white matter) transmit action potentials faster, up to approximately 150 m/s, due to saltatory conduction (Figure 2). In saltatory conduction, ion channels are only necessary between regions of myelination (Nodes of Ranvier); thus, the signal dissipates quickly under regions of myelination to each node and is re-established there, until the signal reaches the terminus. On the other hand, in unmyelinated axons (gray matter), conduction is continuous. This means signal transmission is relatively slow because the voltage gated ion pumps must be present, and used, on all regions of the axonal membrane. Continuous conduction speeds rarely reach 10 m/s. Nerve conduction velocities, which are calculated as the result of the transmission of hundreds or thousands of action potentials firing along the axons of neurons inside nerves, may be affected by the health of the nervous system, age, sex, and even temperature.
Electromyograms (EMGs) are a non-invasive way to determine nerve conduction velocities in the clinical setting. In one type of nerve conduction velocity test (NCV) an EMG recording electrode detects the contraction of a muscle in response to direct neural stimulation by a stimulating electrode placed upon the skin. The time from the onset of stimulus (firing of the stimulating electrode) until muscle contraction is recorded. The stimulating electrode is then moved and the measurement is taken again. From this information, one can determine the nerve conduction velocity. Nerve conduction velocities inform us about the relative health of peripheral nerves. Damage to nerves or diseases that cause demyelination (such as multiple sclerosis – MS) or nerve degeneration (like amyotropic lateral sclerosis – ALS) are primary concerns when conduction speeds are undetectable or below the normal range. The peripheral nerves of the arms generally have conduction velocities between 50-60 m/s.
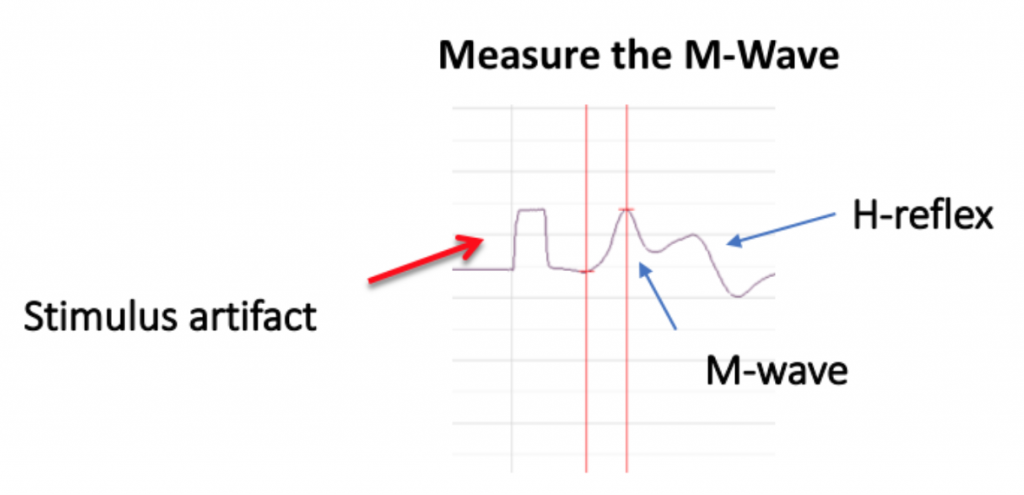
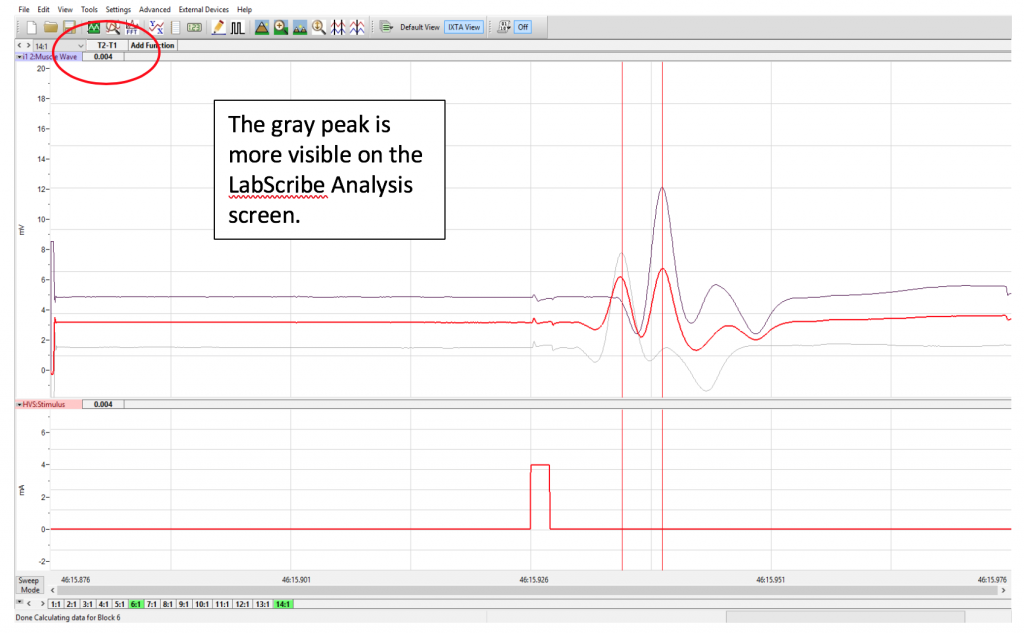
Because there are both sensory and motor fibers in the Ulnar nerve, both will be stimulated upon application of stimulating electrodes to the skin. Activation of sensory fibers will initiate the “H-reflex,” for which the pathway includes the sensory neurons, integrating center (spinal cord), and motor neurons. This is much like we saw last week in the muscle spindle reflex experiments. Application of current to the ulnar nerve also results in an M-wave, an EMG wave due to direct activation of the motor neurons, which will quickly produce a muscle response. In this lab, we will take measurements from the M-waves, only. Note that passive transmission of current in body tissues from the stimulating electodes also produces a stimulus artifact on the EMG. Please see the figure below (Figure 3).
This week, we will perform one experiment (lab exercise 1) and one clinical evaluation (lab exercise 2). The experiment will determine the effects of increasing stimulus strength on the EMG. Stimulus (measured in milliamperes, mA) is applied directly to the fibers of the ulnar nerve with stimulating electrodes on the skin. The resulting EMG amplitude is recorded in mV. EMG amplitude measures the strength of contraction by the innervated muscle, which is relative to the number of motor units activated by the stimulus. Recruitment of more motor units (activation of additional motor neurons and their innervated muscle cells) is accomplished by increasing the stimulus. Next, we will perform a nerve conduction study using EMG. Here, we will use the optimal stimulus strength from Experiment 1 to re-stimulate the ulnar nerve. After recording the location of the stimulating electrode on the arm and obtaining the EMG, the stimulator will be moved to a secondary position, re-stimulating the ulnar nerve. Here, we will divide the physical distance between the stimulators (in mm) by the difference in time between the two trials’ M-waves on the EMG. This method of calculating conduction velocity is called the difference method.
Laboratory Methods Highlights
· This week, instead of stimulating a nerve with a reflex hammer via stretch receptors, we will stimulate a nerve directly with stimulating electrodes.
· Please wear short sleeves or sleeves that can be rolled above the elbow, half way up the brachium.
· A stimulating electrode delivers a mild electric shock, which, if sufficiently strong, depolarizes the membranes of neurons, causing them to fire.
· Neurons stimulated with stimulating electrodes behave like other neurons, except that when stimulating the fibers of a nerve, we may provoke action potentials in both sensory and motor fibers at the same time.
· In this lab we will stimulate the ulnar nerve and determine the outcome of increasing stimulus strength (in mAmps).
· Later, we will re-stimulate the ulnar nerve from two different locations along the arm in order to determine the conduction velocity.
Equipment Required: IXTA data acquisition unit, iWire-B3G cable and three EMG lead wires, disposable snap electrodes, HV stimulator lead wires.
Turn on the iWorx hardware box at the switch on the back, and select the Week7_NerveConduction.iwxset file from the P-drive.
3. Clean the areas where the electrodes will be attached with an alcohol pad. Lightly abrade the skin in those areas.

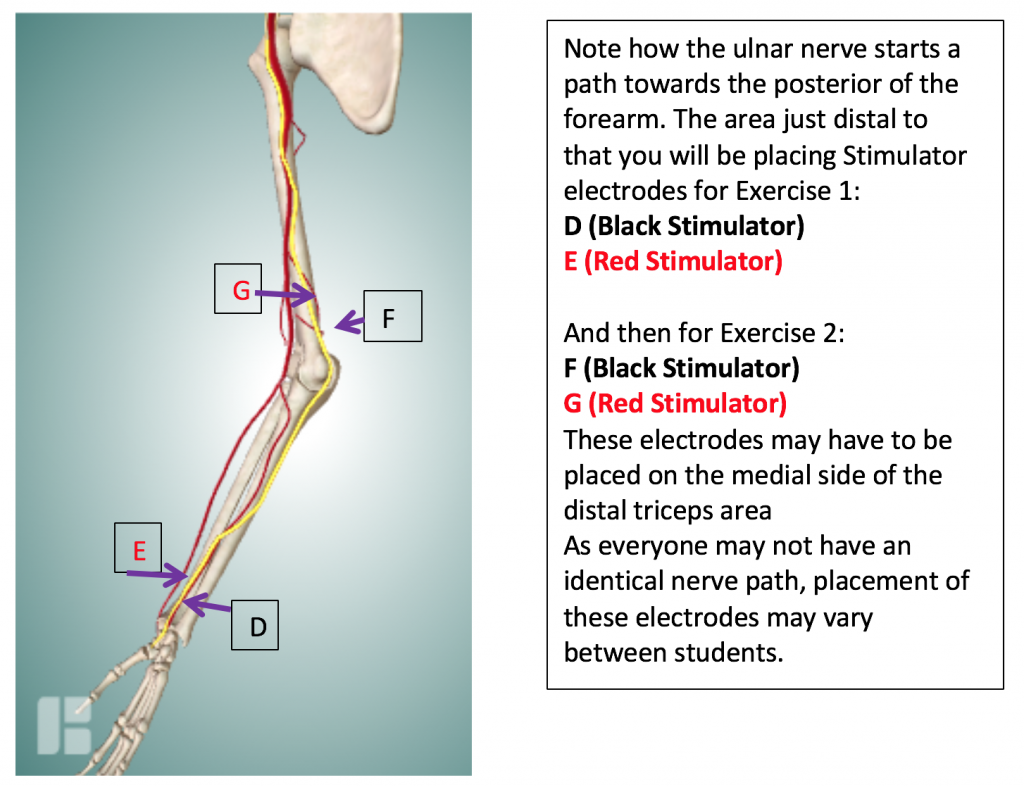
Next, attach the remaining 2 electrodes to the subject’s arm so that they are placed on the pathway of the Ulnar Nerve in the following configuration:
You will deliver mild electric shocks to either yourself or a volunteer experimental subject. The equipment you use to do this is carefully designed to keep the parameters of the electroshocks well within a safe range, and this experiment is safe and fun. Nevertheless, we ask that you place stimulating electrodes on the arms only, and only place electrodes on the same side of the body (never on both arms at the same time).
If you think you may be pregnant or if you suffer from known heart conditions or have an artificial heart pacemaker please do not volunteer as an experimental subject this week.
Objective: To determine the effect of stimulus strength on the response of the innervated abductor digiti minimi muscle.
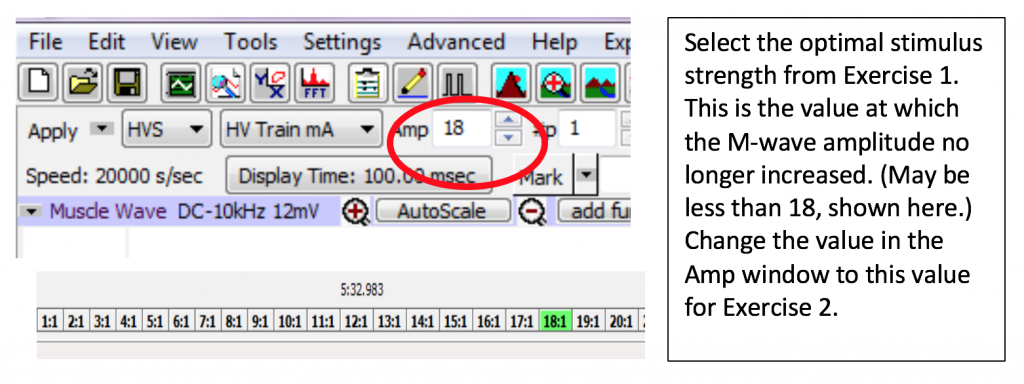
Overview: You will stimulate the abductor digiti minimi motor neuron with increasing amounts of current (measured in milliAmps, read as AMP on LabScribe). You will measure the EMG muscle response each time (in mV) and record the value in your lab report. You will continue until you get three readings in a row with the same value (+/- 0.2 mV). Of those three readings, put a mark next to the lowest current (Amplitude) value on your lab report; you will be using this value for exercise 2.
Continue to increase the output amplitude by an increment of 1 mAMP (1 “AMP” on the LabScribe stimulator button) until you get a muscle twitch and EMG M-wave for the subject.Click apply.
Remember, after each increase in amplitude, you must click “APPLY” before doing another recording!
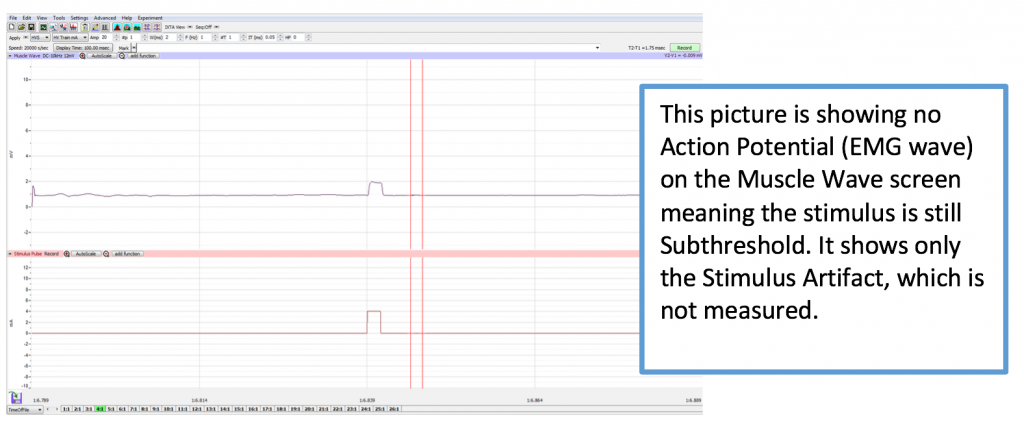
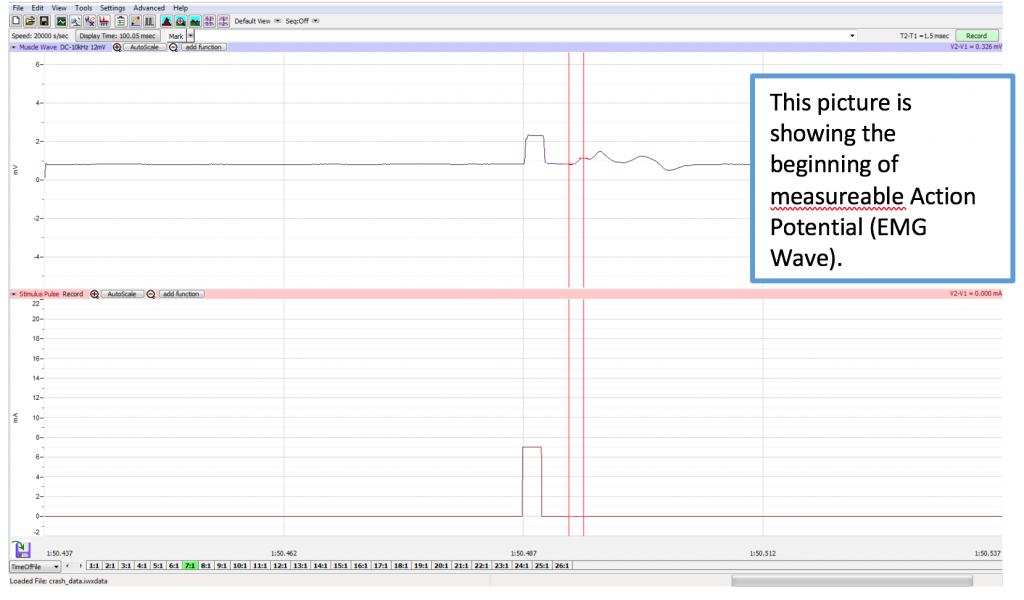
Different sweeps may be selected any time by clicking on the Sweeps list on the bottom of the Main Screen (1:1, 2:1, etc.). If analyzing data on the Main Screen while recording, the sweep you need to analyze is already selected. The Sweeps task bar is shown below.
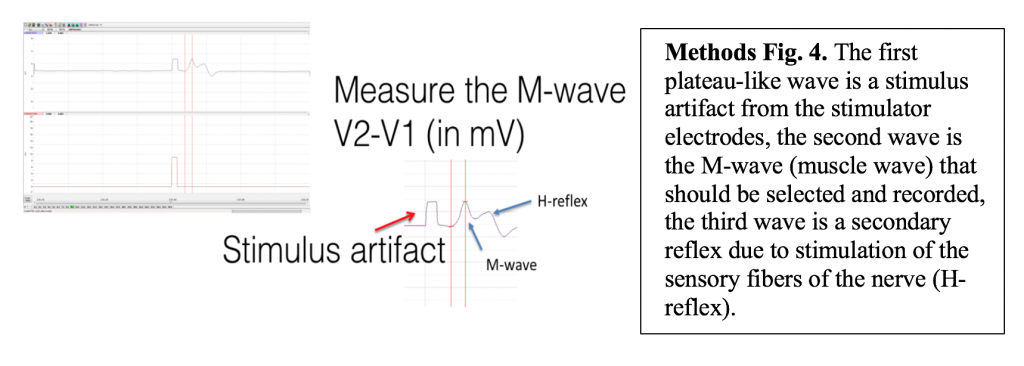
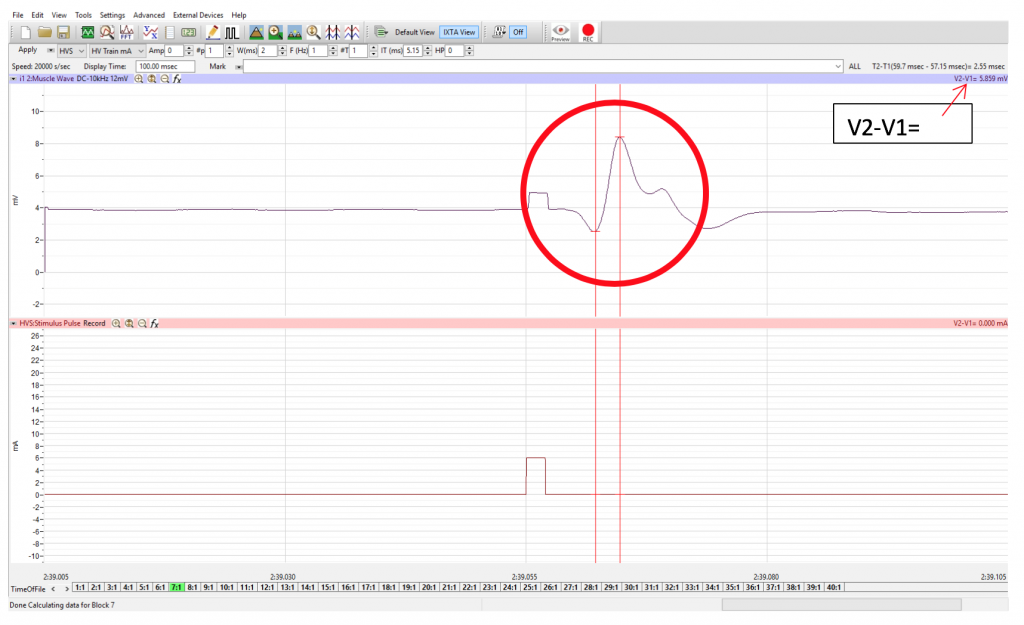
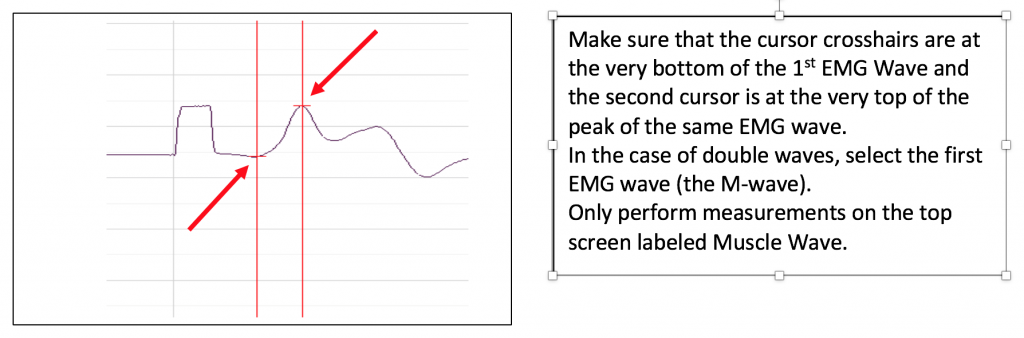
Objective: To measure the conduction velocity (transmission speed) of the ulnar nerve.
You will use the AMP value which excited the maximal response in experiment 1 and stimulate the same abductor digiti minimi motor neuron further up the arm. You will then use that previous experiment and the one you will conduct here to compare the time between innervation and the elicitation of the response for two points on the nerve. You will measure the distance between the stimulation electrodes and calculate the transmission speed in m/s.
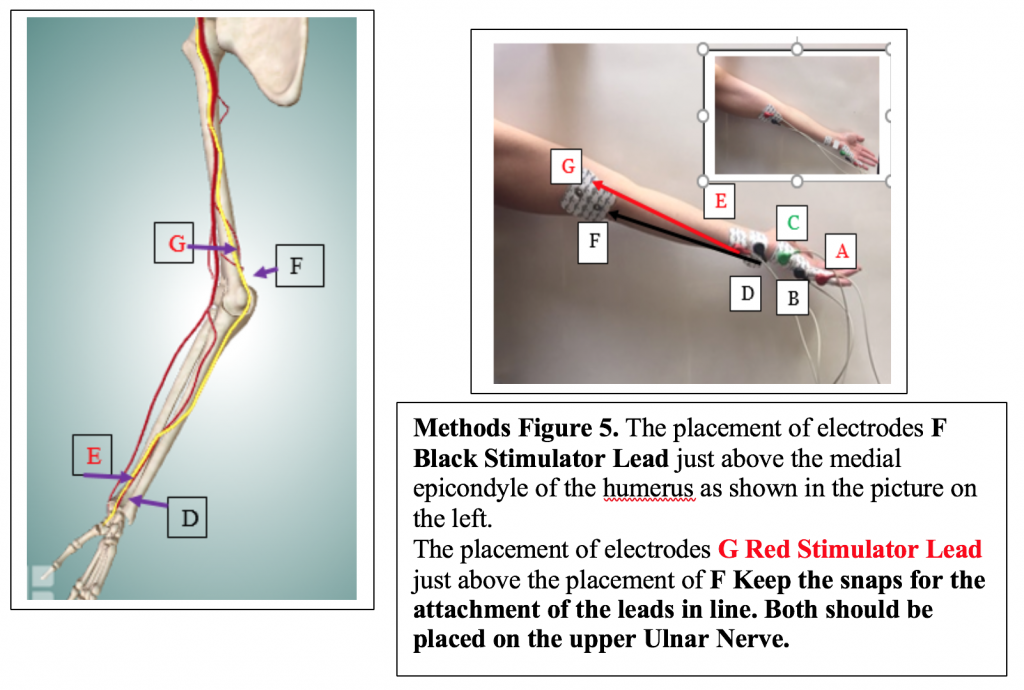
Test the placement by setting the Amplitude to the number that produced the optimal Action Potential previously. Click Apply and Click Record. If you get an Action Potential on the EMG screen proceed to Step 1 below.
If you do not get an Action Potential, reposition F and G electrode pads and test again. Ask your TA for help. Reposition the stimulator electrodes until you get an Action Potential then proceed to step 1 below.
This page titled 2.8: Human Nerve Conduction Velocity (NCV) is shared under a CC BY-SA 4.0 license and was authored, remixed, and/or curated by Karri Haen Whitmer (Iowa State University Digital Press) via source content that was edited to the style and standards of the LibreTexts platform.